把研究感觉信息处理过程作为揭示脑的奥秘的突破口,其中以视觉系统的研究最为突出。在视知觉的研究中已取得了一系列成果:
1、视网膜的光感受器水平:已克隆出视色素蛋白基因;光电换能过程的第二信使是cGMP(Ca2+),黑暗中,cGMP的作用使Na+通道蛋白活化,导致Na+通道开放,Na+持续内流(暗电流),光感受器细胞去极化;光照引起视色素分解,使视盘膜上的GTP结合蛋白分子活化,后者再激活PDE,迅速分解cGMP,引起Na+通道关闭,暗电流骤降,光感受器细胞膜超极化,这样光能就转化为神经电信号。
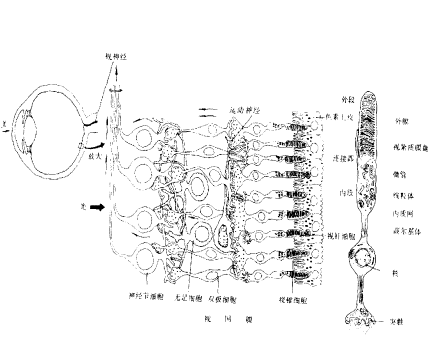
2、视网膜,复杂的信息处理(外周脑),研究相当清楚。视网膜这个两维的、多层次信息处理的最后结果,是经由视网膜神经节细胞以动作电位脉冲调频的方式,传递给脑的。
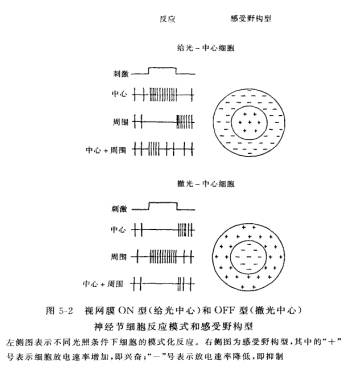
3、感受野(receptor field):视通路中任一神经元都在视网膜(或视野)上有一个代表区域。同心圆拮抗型感受野,包括给光—中心和撤光—中心两类,为心理学马赫带现象提供生理学基础,非同心圆的感受野的细胞对快速运动、运动方向以及某些图形特征产生反应。
4、初级视皮层(纹状皮层),在整个大脑皮层研究最透彻的一部分,面积最大的区域。功能柱:具有相同感受野位置和生理功能的细胞按垂直于皮层表面的柱状结构有序地排列起来。功能柱内细胞具有相同的最优方位、相同的眼优势、相同的最优空间频率。
基本的视觉信息:
1、亮度:可见光的波长范围为380~780nm,亮度是一种外界辐射的物理量在我们视觉中反映出来的心理物理量。亮度定义:光度学计量的一种人的心理物理量,它是在特定的标准条件下,定义一个量值表示辐射量和人平均的光感觉之间的对应关系。人的视知觉对强度和能量的量度,是平均人眼对光辐射量的主观量度。
2、形状:由物体在视觉空间上的亮度分布、颜色分布或运动状态不同而显示出来。对比度(contrast):定义一条亮暗边界的对比度为最大光亮度Lmax与最低亮度Lmin之差除以两者之和,Lmax-Lmin/(Lmax+Lmin)。实验常用的刺激图有光点、光环、光条、光栅、随机点图形,无不涉及图形与背景的对比度测定,因此对比度是一个关键的视觉刺激量。一条视觉上的亮暗边界还有一个重要的属性,即其在二维空间上的朝向——方位,方位选择性已成为公认的形状视觉中最为重要的视觉刺激参数。
3、运动(运动方向和速度):视觉系统对运动目标十分敏感,兔的视网膜中有大量的对运动目标极为敏感的给光-撤光类的W型细胞。
4、颜色:主观感觉,中枢机制尚不清。例如,用适当的红绿兰三种颜色同时刺激,颜色感觉完全消失,只有亮度感觉。在暗视时,只能区别亮度,单变量原理
明视时,三种视锥细胞共同工作,不同波长的光刺激不同的感受系统的相对程度不同,产生颜色的感觉器水平,色觉的三变量性。
5、立体(或深度)视觉:同一物体在左右眼的视网膜象是有微小差别的,这种差别为视觉系统提供了立体视觉(stereopsis)的最基本的信息——视差(disparity )。立体视觉使我们的得到了一个对周围世界的生动而精确的相对的深度分辨。只有在双眼同时运作才有的感觉。
二、视觉的解剖结构
图1是眼球的结构,一束平行光线从眼球的最前方的角膜穿过,经过前房、晶状体、玻璃体,最后聚焦于视网膜上的一点。整个眼球的折光装置类似于一个凸透镜,其中晶状体是可以调节的,根据视物的远近进行调节,光线的强弱可以调节瞳孔的大小。如果视点落在视网膜前,则为近视,反之为远视。一般是通过眼镜来调节折光率,目前有一些治疗近视的技术是处理角膜,改变角膜的折光性。
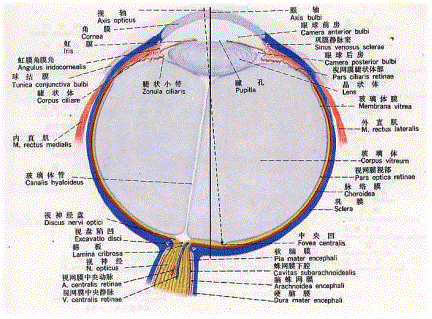
图 1 眼球结构
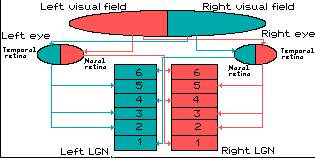
人视网膜可分为以视轴为中心直径约5~6mm的中央区和周边区。中央区又可分为中央凹(fovea)、旁中央凹和远中央凹;周边区也分为近周边区、中周边区和远周边区。 光线落在视网膜上后,被感光细胞接受,产生超极化响应,将光信号转化为电信号。然后通过图2的视觉传导通路传递到脑皮层的视区,形成视觉。视觉传导途径:感受器(视杆和视锥细胞)→第一级神经元(双极细胞)→第二级神经元(节细胞)→视神经→视交叉(鼻侧半纤维交叉,颞侧不交叉)→视束→第三级神经元(外侧膝状体)→视辐射→内囊枕部→枕叶视区。视束中的纤维有部分经上丘臂进入上丘和顶盖前区→组成顶盖脊髓束→参与视、听觉反射。[/align][align=left]视觉传导通路的不同部位损伤所引起的视野的变化:
a.一侧视神经损伤→患侧视野全盲
b.视交叉中央部(交叉纤维)损伤→双侧视野颞侧偏盲
c.视交叉外侧部损伤→患侧视野鼻侧偏盲
d.一侧视束视辐射或视觉中枢损伤→双眼视野对侧同向性偏盲(即患侧视野鼻侧半偏盲和健侧视野颞侧半偏盲)
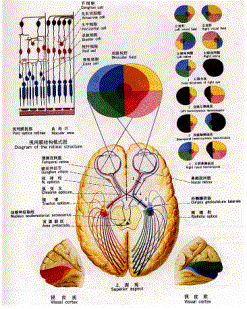 图2 视觉通路
图2 视觉通路